
Dr. Andreas Müller, Düsseldorf
Fledermäuse sind in der Lage, sich in völliger Dunkelheit mittels Echoortung zu orientieren. Manche Forscher, wie Gerhard Neuweiler sprechen lieber von einer „Echoabbildung“, da Fledermäuse nicht nur ihre Umgebung oder ihre Beute orten, sondern sich mithilfe des Ultraschalls sich auch ein genaues Bild machen können. Da der Begriff Echoortung (im englischen echolocation) in der Literatur aber gebräuchlicher ist, soll dieser hier verwendet werden. Dass sich Fledermäuse mit Hilfe des Ultraschalls orientieren, ist noch nicht so lange bekannt. Erst 1938 konnte dieser Zusammenhang sicht- bzw. hörbar gemacht werden.
Geschichte der Echoortung
Im 18. Jahrhundert führte der italienische Bischof und Universalwissenschaftler Lazzaro Spallazani (1729-1799) einen Versuch mit Eulen und Fledermäusen durch. Er holte sich beide Tierarten in sein Studierzimmer und stellte fest, dass sich Eulen weigerten durch den Raum zu fliegen, wenn er alle Kerzen gelöscht hatte. Anders verhielt sich dies mit den Fledermäusen, diese flogen zielsicher durch das Zimmer und berührten auch keinen, der von Spallazani an die Decke gehängten Drähte, an denen kleine Glöckchen befestigt waren. Dann erweiterte er den Versuch, mit nicht nur aus ethischer Sicht, sondern auch mit den heutigen Tierschutzgesetzen unvereinbarer Weise, indem er die Fledermäuse mit glühenden Stricknadeln blendete.
Trotz ihrer Blindheit berührten die Fledermäuse auch in völliger Dunkelheit keinen der Drähte. Erst als Spallazani in die Ohren der Fledermäuse kleine Messingröhrchen klebte und diese mit Wachs verstopfte, flogen die Fledermäuse gegen die Drähte mit den Glöckchen. Wurden die kleinen Messingröhrchen in den Ohren wieder geöffnet, flogen die Fledermäuse problemlos durch den Raum. Da die Fledermäuse allerdings für Spallaziani stumm und lautlos durch die Dunkelheit flogen, blieb für ihn das Geheimnis des Ultraschalls verschlossen. Er postulierte eine Art sechsten Sinn, den die Fledermäuse besitzen sollen. Ein Zeitgenosse Spallazanis, der berühmte Naturforscher Georges Baron de Cuvier (1769-1832) zog aus diesen Experimenten den Schluss, dass es wohl der äußerst feine Tastsinn der Fledermäuse ist, der ihre Orientierung im Dunkeln ermöglicht. Er stellte die These auf, dass Fledermäuse mit ihren Flügeln Luftstauungen wahrnehmen, die bei der Annäherung an ein Objekt entstehen. Das war lange Zeit der Stand der Wissenschaft in Bezug auf die Orientierung der Fledermäuse.
Erst 150 Jahre nach Spallazanis Versuchen endeckten Wissenschaftler mit Hilfe moderner Technik, dass Akustik des Rätsels Lösung ist. Diese Entdeckung gelang zwei Wissenschaftlern unabhängig voneinander, dem amerikanischen Zoologen Donald Redfield Griffin (1915-2003) und dem niederländischen Zoologen Sven Dijkgraaf (1908-1995). Griffin konnte zusammen mit dem Physiker George Washington Pierce (1872-1956) mit einem Hochfrequenzdetektor nachweisen, dass die scheinbar lautlosen Fledermäuse intensive Lautäußerungen im Ultraschallbereich produzierten und er konnte diese auch hörbar machen, was beide 1938 im Journal of Mammalogy publizierten. Etwa zeitgleich stellte Dijgraaf, der ein außergewöhnliches Gehör hatte und so die niedrig-frequenten Laute der Fledermäuse hören konnte fest, dass Fledermäuse Laute zur Orientierung ausstoßen. Hierfür verpasste er Fledermäusen Maulklappen, die sich öffnen und schließen ließen. Bei offener Klappe konnten sie problemlos durch den Raum fliegen, aber bei geschlossener Klappe kamen sie ins Schlingern. Dijkgraaf nannte dieses Konzept Echolokalisation und publizierte dies 1943.
Die Erforschung der Echoortung von Fledermäusen in Deutschland wurde u.a. durch den Zoologen Gerhard Neuweiler (1935-2008), sowie eine Forschungsgruppe der Abteilung Biologie II an der Ludwig-Maximilian-Universität München weitergeführt.
Ultraschall
Als Ultraschall bezeichnet man den Schall mit Frequenzen oberhalb des Hörfrequenzbereichs des Menschen. Er umfasst Frequenzen ab 20 kHz bis 10 GHz; Schall oberhalb dieser Grenze wird als Hyperschall bezeichnet. Der Schall mit Frequenzen unterhalb der Hörgrenze des Menschen, d. h. <16 Hz, wird Infraschall genannt. Der Hörbereich des Menschen liegt also zwischen 16 bis 20 Hz und 16 bis 20 kHz, wobei Kinder ein deutlich besseres Gehör im höheren Frequenzbereich haben und so die tieferen Töne der Fledermäuse hören können.
Das Prinzip Echoortung
Das Echo-Abbildungssystem der Fledermäuse besteht aus einem Sender, dem Kehlkopf, der die Laute erzeugt und einem Empfänger, den Ohren und ist ein aktives Orientierungssystem, da die Laute vom Tier selbst erzeugt werden. Allerdings hat diese Art der Orientierung auch mehrere Nachteile. Die akustische Abbildung kostet Energie, die die Fledermaus in Form von Nahrung wieder für sich gewinnen muss. Ein weiterer Nachteil ist, dass anders als bei der optischen Wahrnehmung, bei der die Umgebung ständig abgebildet wird, bei der akustischen Abbildung diese nur stroboskopisch abgebildet wird, wenn das Tier einen Laut aussendet. Dies kann man sich vorstellen, wie wenn früher in Diskotheken das Licht schnell flackerte, wodurch eine Art Zeitlupeneffekt entsteht. Der dritte Nachteil ist das kleine Schallfeld, verglichen mit dem großen Gesichtsfeld eines Säugetiers. Diese Schallkeule, die von der Fledermaus ausgesendet wird ist eingeengt und auf die Flugrichtung ausgerichtet, d.h. Alles, was außerhalb dieses Schallfeldes liegt, wird nicht erfasst. Nur wenn Fledermäuse ihre Umgebung mit ständigen Ortungslauten abtasten, entsteht ein weiträumiges Echobild der Umgebung. Der vierte Nachteil ist die begrenzte Reichweite der Echoabbildung, meist zwischen 20 und 60 Metern, wobei tiefere Frequenzen eine höhere Reichweite haben. Ein letzter Nachteil ist die limitierte Auflösung, wobei hohe Frequenzen eine bessere Strukturauflösung erzielen als niedrige. Die Fledermaus muss also einen Kompromiss finden zwischen großer Reichweite bei tieferer und einer besseren Auflösung bei höheren Ortungsfrequenzen.
Evolution der Echoortung
Unter den Landsäugetieren haben nur Fledermäuse ein Echoortungssystem entwickelt. Die meisten Kleinsäuger, wie z.B. Spitzmäuse, können zwar Ultraschall gut hören, aber sich nicht mit dessen Hilfe orientieren.
Arten der Unterordnung Yangochiroptera (Fledermäuse im eigentlichen Sinne) und die Hufeisennasen aus der Unterordnung Yinpterochiroptera (Flughunde und Hufeisennasen) besitzen in erster Linie Echoortung. Bei fast allen Flughunden fehlt sie, nur die Gattung Rousettus, zu der der einzige in Europa auf Zypern vorkommende Flughund, der Nilflughund (Rousettus aegyptiacus) zählt, der Höhlen bewohnt und sich mit Hilfe der Echoortung in diesen orientieren kann. Da Flughunde in erste Linie dämmerungs- oder nachtaktiv sind, sich von Früchten bzw. Samen ernähren und große Augen besitzen, reicht bei ihnen das Restlicht in der Dämmerung und in der Nacht zur Orientierung und zur Nahrungsfindung aus. Zusätzlich können sich Flughunde durch Schnalz- oder Klicklaute mit der Zunge orientieren.
Da Echoortung in beiden Unterordnungen der Fledertiere vorkommt, wird in einer aktuellen Hypothese diskutiert, ob sie sich zweimal unabhängig entwickelt hat. Außerdem existieren weitere ältere Hypothesen darüber, was sich zuerst entwickelt hat, die Echoortung, der Flug oder beides zusammen (Tandem-Hypothese).
Zur Entstehung der Echoortung, gibt es zwei Hypothesen: Entweder sie hat sich aus der Notwendigkeit entwickelt, sich in Höhlen zu orientieren, da höhlenbewohnende Fledermäuse und Flughunde diese besitzen oder die Echoortung entstand aus der Notwendigkeit Insekten beim nächtlichen Flug als Beute zu erkennen, da das schwache Restlicht in der Nacht nicht mehr ausgereicht hat. Der Selektionsdruck, der von der Insektenjagd ausging, dürfte dabei stärker gewesen sein als der durch die Besiedlung von Höhlen.
Ortungslaute, ihre Erzeugung und ihr Empfang
Die Ortungslaute der Fledermäuse werden im Kehlkopf erzeugt, sind im Unterschied zu Kommunikationslauten, wie z. B. den Soziallauten der Fledermäuse, kurz und dauern nur wenige Millisekunden (ms) an. Man unterscheidet drei Signalelemente:
a) Frequenzmodulierte Abwärtslaute (FM ab): Dies sind die häufigsten Ortungssignale, die bei einer hohen Frequenz beginnen und kontinuierlich zu niedrigeren Frequenzen abfallen.
b) Reine Töne (CF=constant frequency): Dies sind Töne bei einer konstanten Frequenz, die vor allem als Suchsignale ausgestoßen werden und besonders bei Hufeisennasen (Rhinolophus) häufig sind.
c) Frequenzmodulierte Aufwärtslaute (FM auf): Dem konstanten Frequenzteil folgt ein frequenzmodulierets Element voraus, der FM Aufwärtslaut, was ebenfalls häufig bei Hufeisennasen auftritt.
Eine punktförmige Schallkeule strahlt Schallenergie in Form einer Kugelwelle ab. Dabei wirkt das geöffnete Maul der Fledermaus, bzw. bei Hufeisennasen die Nasenlöcher als Schalltrichter, der die Hauptenergie des Lautes in Flugrichtung bündelt, wodurch die Schallkeule entsteht. Die beiden Ohren empfangen die Ortungslaute und dienen als Schallrezeptoren. Dabei sind die, bei Fledermäusen dreh- und neigbaren Ohrmuscheln bewegliche Richtantennen und der Schall wird durch den Gehörgang weiter geleitet bis zum Trommelfell. Da Fledermausarten unterschiedliche Ohrmuscheln haben, haben diese auch verschiedene akustische Eigenschaften. Die Ohrmuscheln der meisten Echo-Orter sind nicht auffallend groß, da sie auf Ultraschall mit kurzen Wellenlängen abgestimmt sind. Andere Arten, die ihre Beute vom Boden ablesen, wie beispielsweise Langohren (Plecotus) haben dagegen große Ohrmuscheln, die auch auf das Hören des Raschelns der Beute im Laub auf dem Boden spezialisiert sind. Der Tragus (Ohrdeckel), eine steife Hautfalte in der Ohrmuschelöffnung, spielt wahrscheinlich bei der vertikalen Schalllokalisation eine Rolle. Bei den Hufeisennasen fehlt dieser Tragus, sie haben einen sogenannten Antitragus, eine horizontale Hautfalte am Boden der Ohrmuschelöffnung. Das Mittelohr der Fledermäuse besteht aus der Paukenhöhle, die vom Trommelfell begrenzt wird. Drei Gehörknöchelchen, der Hammer, der Amboss und der Steigbügel durchqueren die Paukenhöhle. Die eintretende Schallenergie versetzt das Trommelfell in Schwingungen, die über die Gehörknöchelchen auf das Innenohr übertragen werden.
Das Mittelohr der Fledermäuse ist besonders an die Übertragung hoher Frequenzen, wie sie bei Ultraschallauten entstehen angepasst. Das Innenohr von Fledermäusen löst die Schallfrequenzen auf Zehntelprozent genau auf. Die drei Flüssigkeitsräume des Innenohrs sind in eine knöcherne Kapsel eingebettet und wie eine Schnecke spiralig gewunden, deshalb auch ihr Name „Cochlea“. Haarzellreihen übertragen die Schallenergie auf kurze, bewegliche Härchen der Sinneszellen, die sog. Stereozilien. Jede Haarzelle trägt ein Büschel Stereozilien, von denen Nervenzellen dann die Hörinformation übernehmen.
Ortungsleistungen
Die Echoortung wird in erster Linie zur Detektion und Lokalisation genutzt. Sie dient nicht, wie beispielsweise bei Vögeln, der Kommunikation der Tiere untereinander. Eine Fledermaus sendet im Suchflug nach Beute etwa 4 – 12 Ortungslaute pro Sekunde aus, die oft mehr als 10 Millisekunden lang sind. Da die Pausen zwischen den Lauten jedoch wesentlich länger sind als die Laute, bleiben etwa 4/5 der Flugzeit ohne Echoinformation. Entdeckt die Fledermaus ihr Ziel, werden Laute in rascher Folge ausgestoßen und mit immer kleineren Pausen. Bei der Verfolgung der Beute steigen die Wiederholungsraten auf 40 – 50 Laute pro Sekunde. Kurz bevor die Beute gepackt wird, sendet die Fledermaus eine Folge von 10 – 25 kurzen Lauten mit minimalen Pausen aus, den sogenannten „final buzz“. Der ganze Vorgang von Detektion der Beute bis zum „final Buzz“ dauert meist weniger als eine Sekunde.
Die Detektion einer Beute im freien Luftraum ist für eine Fledermaus kein Problem, da das von ihr aufgenommene Echo in der Regel ein fliegendes Insekt bedeutet. Die Beute wird lokalisiert und die Entfernung zur Beute gemessen.
Vertikale und horizontale Echorichtungen können Fledermäuse auf 2 – 5° genau erkennen, allerdings ist der Hörraum, aus dem das Echo zu erwarten ist, durch die Schallkeule eingeschränkt. Die Ohrmuscheln tragen zur Richtungsfindung des Echos bei, da sie beweglich sind und Schallwellen abschirmen, die auf ihre Rückseite treffen. Für ein bodenbewohnendes Tier ist die Bestimmung der vertikalen Schallrichtung nicht so entscheidend, wie für fliegende Tiere, die ihre Beute in der Luft verfolgen. Hierbei hilft der Tragus der Fledermäuse, die häufig breitbandige Ortungssignale aussenden. Für die Bestimmung der horizontalen Schallrichtung einer Quelle eignen sich die Reizdifferenz zwischen linkem und rechtem Ohr der Fledermaus, d.h. mit welchem Ohr die Beute zuerst wahrgenommen wird.
Die kleinste, für Fledermäuse akustisch wahrnehmbare Objektgröße liegt knapp unter 1 mm. In Versuchen wurden 2 bis 4 mm große Fruchtfliegen aus einer Entfernung von etwa 35 cm von Fledermäusen entdeckt und gefangen. Verschiedene Fledermausarten konnten in Versuchen Drahthindernissen bis zu einem Durchmesser von 0,06 bis 0,10 mm ausweichen. In anderen Versuchen wurde gezeigt, dass Fledermäuse Platten mit 8 mm tiefen Bohrlöchern von Platten mit 7 mm tiefen Bohrlöchern unterscheiden konnten, was darauf hindeutet, dass Fledermäuse auch die Oberflächenstruktur ihrer Umgebung wahrnehmen können. Dies zeigten auch Untersuchungen von Wissenschaftlern der Ludwig-Maximilian-Universität München, die belegen konnten, dass Echos Informationen enthalten, die es den Tieren ermöglichen, verschieden strukturierte Oberflächen voneinander zu unterscheiden. So sticht zum Beispiel für Fledermäuse eine zappelnde Beute selbst auf einer bewegten Wasseroberfläche akustisch heraus.
Dabei wirkt die glatte Wasseroberfläche akustisch wie ein Spiegel und der Schall wird im gleichen Winkel, in dem er auf die Wasseroberfläche trifft, auch wieder reflektiert – hauptsächlich von der Fledermaus weg, da sie schräg nach unten-vorne auf das Wasser ruft. Ein treibendes Insekt reflektiert den Schall jedoch direkt zur Fledermaus zurück, als ein einzelnes Beuteecho inmitten von Stille. Ist die Wasseroberfläche aber nicht glatt, sondern hat viele Wellen, können Fledermäuse ihre Beute am besten orten, wenn möglichst viele Wellen auf enger Fläche sind, die Oberfläche also „geriffelt“ ist. Die Forscher deuteten das Ergebnis so, dass sanfte Hintergrundwellen, wie sie durch Wind entstehen, für die Fledermäuse praktisch unsichtbar sind, während eine in den Wellen zappelnde Beute viele Wellen erzeugt und so gut von der Fledermaus detektiert werden kann.
Biotopsanpassungen
Fledermäuse jagen bevorzugt in Arealen. An diese Jagdbiotope haben sie nicht nur ihren Flugstil und ihre Flügelform angepasst, sondern auch ihr Ortungssystem. Allerdings beschränken Fledermäuse sich nicht nur auf das bevorzugte Jagdgebiet. sie weichen auch auf andere Biotope aus, beispielsweise in Jahreszeiten mit reduziertem Beuteangebot oder wenn in Schlechtwetterperioden die Insekten in tieferen Luftschichten fliegen.
Die Echoortung eignet sich besonders für die Insektenjagd im freien Luftraum. Die Jagd im Geäst der Bäume verspricht zwar ein reicheres Insektenangebot, allerdings kann das Echo der kleinen Beute durch die Echos der größeren Vegetation im Hintergrund überlagert werden.
Alle Fledermäuse, die im freien Luftraum jagen, benutzen bei der Beutesuche lange, niedrigfrequente Ortungslaute. Haben sie eine Beute gefunden, wird der Ortungslaut in ein kurzes (1 bis 5 ms), frequenzmoduliertes, breitbandiges Signal umgewandelt, das gut zur Zielentfernungsmessung geeignet ist. Dabei korreliert die größere Reichweite der niederfrequenten Ortungslaute mit einer schlechteren Detektion keiner Insekten.
Fledermausarten, die am Blattwerk oder über den Boden jagen, haben mit dem „Echorauschen“ durch den Hintergrund zu kämpfen. Sie müssen das Echo eines Insektes aus der Vielzahl der Echos, die der Hintergrund zurückwirft, herausfiltern. Das Große Mausohr (Myotis myotis) fängt im Wald bodenbewohnende Insekten – Laufkäfer, aber auch Spinnen und Tausendfüßler. Dabei sendet es gewöhnliche, frequenzmodulierte Signale aus und fängt das Insekt mit dem, für die Echoabbildung im Flug typischen „final Buzz“. Wird die Beute jedoch im Abstand von nur wenigen Zentimetern an einer Wand präsentiert, verstummt das Mausohr und läuft die Wand ab, um die Beute zu ertasten.
Hufeisennasen, die ihre Beute bevorzugt im Flug zwischen dichter Vegetation jagen, fliegen die ersten Abendstunden um Büsche und Bäume. Später betätigen sich als Lauer-Jäger, die im Geäst der Bäume sitzen, sich um die eigene Achse drehen und dabei die Umgebung per Ultraschall absuchen. Trifft der Ortungslaut auf ein fliegendes Insekt, holt sich die Hufeisennase dieses und kehrt dann zu ihrem Ast zurück. Der Ortungslaut besteht aus einem lauten, 40 bis 60 ms langen Reinton mit hoher Frequenz von 72 bis 85 kHz und mit diesem erkennen Hufeisennasen besonders flügelschlagende Insekten.
Dabei müsste diese Detektion flügelschlagender Insekten zusammenbrechen, sobald die Fledermaus selber losfliegt, da entsprechend der eigenen Fluggeschwindigkeit, das gesamte Tonecho eine Frequenzerhöhung durch einen zweifachen Dopplereffekt, der jeweils am fliegenden Sender (Den Nasenlöchern der Hufeisennase) und am fliegenden Empfänger (den Ohren) entsteht.
Der Dopplereffekt besagt, dass wenn sich eine Geräuschquelle auf einen Hörer zu bewegt, werden pro Zeiteinheit mehr Schallwellen empfangen, als ausgesandt wurden, d.h. die Frequenz des Geräusches wird höher. Entfernt sich die Geräuschquelle, so tritt der gegenteilige Effekt ein, die Frequenzen werden tiefer. Dies kann man am Beispiel der Sirene eines Krankenwagens verdeutlichen, kommt der Wagen auf einen zu, ist der Ton höher, fährt er von einem weg, wird der Ton der Sirene tiefer.
Die Fledermäuse begegnen diesem Problem mit einer Dopplereffekt-Kompensation – d.h. sobald das zuletzt gehörte Geräusch eine bestimmte Sollfrequenz übersteigt, werden die als nächstes ausgesendeten Ortungslaute in ihrer Frequenz um einen entsprechenden Betrag abgesenkt, wodurch die durch die eigene Fluggeschwindigkeit induzierten Frequenzverschiebungen des gesamten Echosignals kompensiert werden.
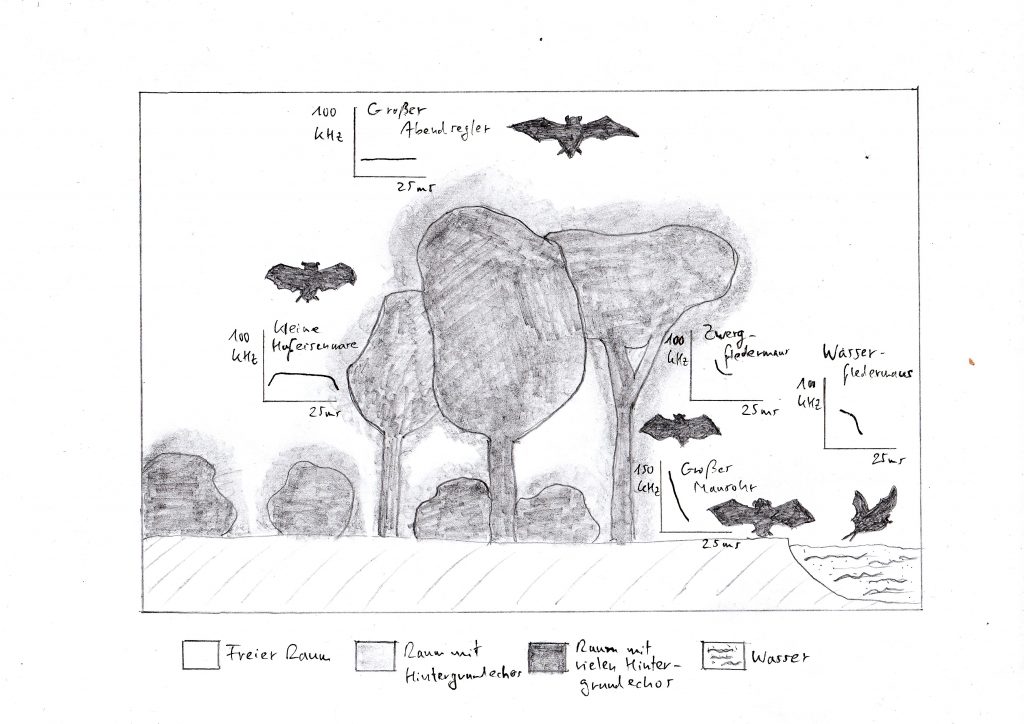
Heimische Fledermäuse in verschiedenen Biotopen und Diagramme der charakteristischen Ortungsrufe – die Silhouetten sind nicht im gleichen Maßstab gezeichnet (verändert nach Schnitzler und Kalko 2001)
Aber genug der Physik, kommen wir zur passiven Ortung. Fledermäuse, die ihre Beute von einem Untergrund fangen, können sich nicht auf die Detektion eines Flügelschlages verlassen. Sie hören ihre Beute daher nur, wenn diese Geräusche, z.B. durch Rascheln im Laub macht.
Gegenmaßnahmen der Beute
Viele Nachtschmetterlinge haben wirksame Abwehrmechanismen zum Schutz vor der Echoortung durch Fledermäuse entwickelt. Sie können die Ultraschalllaute hören und reagieren mit Ausweichmanövern im Zick-Zack-Kurs oder legen ihre Flügel an und lassen sich fallen. Auf Blättern sitzende Schmetterlinge verharren starr, sobald ein Ortungssignal sie erfasst. Einige Nachtfalter aus der Familie der Noctuiden senden ihrerseits Ultraschallsignale, in Form von Klicklauten aus und verwirren so Fledermäuse bei der Jagd, die diese Laute für Warnsignal einer nicht schmackhaften Beute halten. Zerstört man den Klickmechanismus dieser Schmetterlinge, werden sie wieder von den Fledermäusen gejagt und verspeist.
Fledermaus-Detektoren
Da Fledermäuse im Ultraschallbereich rufen, den wir Menschen nicht hören können, müssen wir auf Hilfsmittel zurückgreifen, wie die Bat-Detektoren. Diese Geräte gibt es schon als einfache Version zum Zusammenstecken für etwa 25 € oder auch als elektronischen Bausatz für Bastler für 35 bis 40 €. Andere, auch für Laien gut zu verwendende Geräte, die die Frequenz und die Ausschlaghöhe der Frequenz im Display zeigen, kosten bis zu 160 bis 180 €. Die teuren Geräte, bei denen man die Sonogramme im Display sehen und die man auf SD-Karten speichern kann oder die auf ein Mobiltelefon gesteckt werden können, liegen bei 230 bis 300 €. Im Internet gibt es auch einige gute, allgemein verständliche Publikationen für den Umgang mit Bat-Detektoren zum Nachweis von Fledermäusen, etwa Weid 1988, Zingg 1990, Pfalzer 2007 oder Neuweiler 1990 (siehe unten).
Quellen
Baier A. Leonie, Lutz Wiegrebe and Holger R. Goerlitz (2019): Echo-Imaging Exploits an Environmental High-Pass Filter to Access Spatial Information with a Non-Spatial Sensor. iScience 14: 335–344
Dijkgraaf Sven (1943): Over een merkwaardige functie van den gehoorzin bij vleermuizen. Verslagen Nederlandsche Akademie van Wetenschappen Afdeeling Naturkunde 52: 622–27.
Dijkgraaf Sven (1946): Die Sinneswelt der Fledermäuse. Experientia 2: 438-448.
Neuweiler Gerhard (1993): Biologie der Fledermäuse. Georg Thieme Verlag, Stuttgart.
Neuweiler Gerhard (1990): Echoortende Fledermäuse. Biologie in unserer Zeit 20(3): 169-176
Pfalzer Guido (2007): Verwechslungsmöglichkeiten bei der akustischen Artbestimmung von Fledermäusen anhand ihrer Ortungs- und Sozialrufe. Nyctalus 12(1): 3-14
Pierce George W. and Donald R. Griffin (1938): Experimental determination of supersonic notes emitted by bats. Journal of Mammalogy 19(4): 454-455.
Schnitzler Hans-Ulrich and Elisabeth K. Kalko (2001): Echolocation by insect-eating bats. BioScience 51(7): 557-569
Weid Roland (1988): Bestimmungshilfe für das Erkennen europäischer Fledermäuse insbesondere anhand der Ortungsrufe. Schriftenreihen Bayerisches Landesamt für Umweltschutz 81: 63-72
Zingg Peter E. (1990): Akustische Artidentifikation von Fledermäusen (Marnrnalia: Chiroptera) in der Schweiz. Revue suisse Zool. 97(2): 263-294
© Text und Zeichnung Dr. Andreas Müller, Düsseldorf